 CUBE ChatShaala Meeting Summary (December 12, 2025)
CUBE ChatShaala Meeting Summary (December 12, 2025)
Overview
This session of CUBE ChatShaala convened to review and discuss ongoing student research and fundamental concepts related to seed biology, specifically focusing on seed dormancy and germination kinetics. The discussion was structured around two key areas: the adaptive nature of dormancy in Cardamine species and the comparative germination rates of model seeds.

I. The Temporal Landscape of Seed Dormancy in Cardamine:
The primary focus under the “Seed Dormancy” theme was the genus Cardamine. A comparative timeline demonstrated the species-specific nature of this crucial survival strategy. The analysis highlighted that dormancy for certain species, such as Cardamine hirsuta, is a protracted phase spanning from January to approximately May, with germination commencing in June. This extended period suggests a requirement for specific environmental conditioning, such as prolonged cold or vernalization, to break dormancy. Other species, including Cardamine flexuosa, exhibited a perceptibly shorter dormancy period, emphasizing that even within a single genus, the temporal requirements for reproductive readiness can vary significantly due to localized ecological pressures. The overall principle reinforced was the understanding of dormancy as an essential adaptive trait for maximizing the chances of seedling survival.
II. Comparative Germination Kinetics:
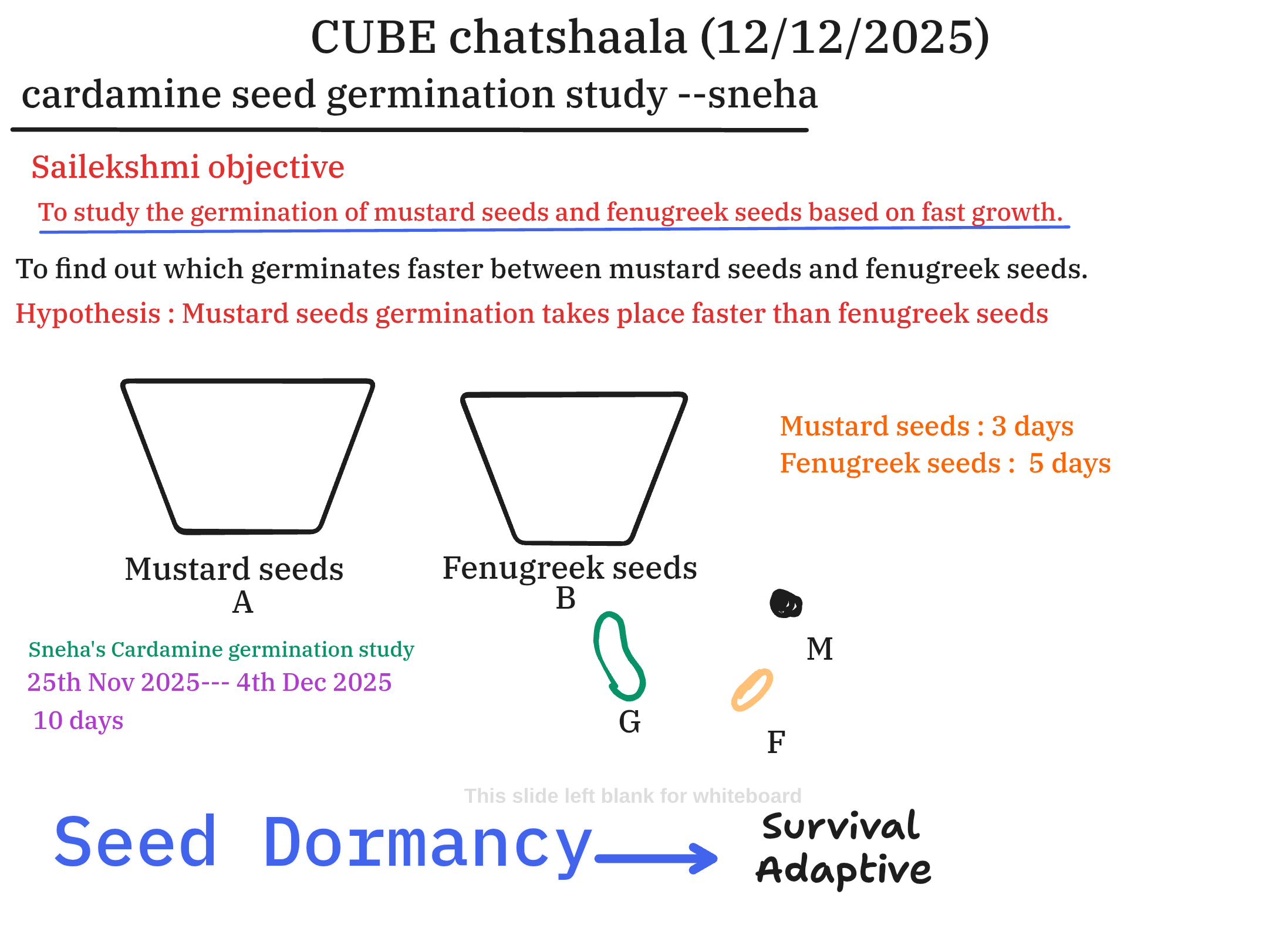
The session included a presentation of two distinct germination studies. Sailekshmi’s work centered on a straightforward comparative experiment between common garden seeds—mustard and fenugreek. The stated objective was to determine which seed germinates faster. The results strongly supported the initial hypothesis: mustard seeds completed germination in approximately 3 days, whereas fenugreek seeds required 5 days. This 40% reduction in germination time for mustard provides a clear, quantitative example of differential germination kinetics. Furthermore, the session acknowledged Sneha’s preliminary 10-day Cardamine germination study, conducted between November 25th and December 4th, 2025, providing a valuable link between the conceptual timeline of dormancy and direct experimental observation.
 Provocative Queries for the General Audience
Provocative Queries for the General Audience
-
The Cardamine timeline shows dormancy from January to May, followed by June germination. Yet, one of the studies was conducted in late November. If dormancy is an adaptive strategy for survival, what specific signal in the ecosystem must be absent for the plant to choose to wait five months, and why do some species wait shorter periods than others? Are we observing a default state of primary dormancy or the variable phenomenon of secondary dormancy based on local environment? What environmental cue is the true “master key” for the successful June germination?
-
Sailekshmi’s study demonstrated that mustard seeds germinate a full two days faster than fenugreek seeds. In a carefully controlled lab setting, this is a significant finding. But in the highly competitive, volatile environment of the wild, where water, light, and nutrients are scarce, does a mere 48-hour head start truly provide a statistically significant advantage for species dominance and survival, or is the post-germination growth rate the ultimate determinant?
 My Learnings, TINKE Moments, Gaps, and Misconceptions
My Learnings, TINKE Moments, Gaps, and Misconceptions
What I Have Learned
I have learned that seed dormancy is not a monolithic trait but a highly nuanced, species-specific temporal adaptation, as evidenced by the distinct dormancy periods of Cardamine hirsuta versus Cardamine flexuosa. I now appreciate that fundamental comparative studies, such as the mustard vs. fenugreek experiment, serve as excellent models for demonstrating that minute differences in germination kinetics (a 2-day variance) can be reliably quantified and linked back to evolutionary adaptive pressures. Crucially, the concept that Seed Dormancy is directly an Adaptive Strategy for Survival has been strongly reinforced.
TINKE Moments (This I Never Knew)
- Species-Specific Calibration: The significant variation in dormancy length among Cardamine species indicates a high degree of evolutionary calibration to specific micro-environments. This suggests that a one-size-fits-all approach to breaking dormancy is insufficient and that the ecological niche is key.
- Quantifying Adaptability: The clear-cut 3-day (mustard) vs. 5-day (fenugreek) result is a powerful insight. It immediately validates the hypothesis and provides a measurable baseline for discussing the relative fitness of different seeds to quickly exploit available resources.
- The Cardamine Complexity: The complexity of the Cardamine life cycle, as inferred from the provided research paper (doi:10.1111/nph.12782 on seed release mechanisms), highlights that the dormancy phase is only one part of a multi-stage adaptive process that includes dispersal and successful establishment.
Gaps and Misconceptions
Gaps Identified:
-
Missing Data from Sneha’s Study: The most significant gap is the lack of explicit, documented results from Sneha’s 10-day Cardamine germination trial (25th Nov - 4th Dec). This data is critical for linking the conceptual Jan-May timeline to current experimental observations.
-
Environmental Context for Dormancy Breaking: The specific environmental cues (e.g., minimum accumulated chill hours, critical light spectrum, or minimum moisture potential) required to break dormancy for the unstudied Cardamine species (bulbosa, lyrata, impatiens) were not detailed.
Potential Misconceptions:
- Primary vs. Secondary Dormancy: The presentation of a Jan-May dormancy timeline may lead to a misconception that this is a fixed, obligate (primary) dormancy. The November study, however, suggests that germination may be possible outside the typical window, raising the question of whether the seeds exhibit conditional or secondary dormancy, where environmental factors could override the typical annual cycle.